官方网站")
NeuroFUS刺激人脑论文分享: 人体经颅超声刺激揭示前额叶-前壳核回路在抑制反应中的因果作用
2022年8月16日来自日本的研究团队在《Cell Reports》(影响因子9.995/Q1)上发表了题为:A causal role of anterior prefrontal-putamen circuit for response inhibition revealed by transcranial ultrasound stimulation in humans的文章,利用NeuroFUS经颅超声刺激(NeuroFUS Pro with TPO-203 & CTX-500)分别刺激了人类初级运动皮层(M1),右半球基底神经节的三个区域(STN、前壳核和后壳核),前额叶皮层中的两个区域(aIFC 和 MFC),结合任务及MRI影像分析表明:“右前壳核和右前 IFC 对反应抑制有因果关系,并表明间接通路中的前 IFC-前壳核回路是反应抑制的重要途径。”本文示范了NeuroFUS经颅聚焦刺激系统用于非侵入性深部脑区精确刺激,也提供了一个可供参考的经颅超声刺激用于高分辨率Brain Mapping的研究范式。
经颅超声刺激 (TUS/tFUS)是一项革命性的非侵入性神经调控技术,“TUS可以无创地刺激大脑深层结构,空间精度高达几毫米”。与其他无创神经调控方法相比,tFUS/TUS提供了前所未有的空间分辨率和精度控制,还可以靶向更深的大脑区域,实现无痛无创非侵入性的精准深部脑刺激,有望为各种神经系统疾病提供新的治疗方法,从慢性疼痛和记忆丧失到心理障碍和失眠。经颅聚焦超声刺激 (TUS/tFUS) 产生的效果可以激活或者抑制特定的大脑区域,因此它有望成为高分辨率Brain Mapping的革命性工具。
论文原文参见DOI://doi.org/10.1016/j.celrep.2022.111197
主要部分翻译整理如下:
人体经颅超声刺激揭示前额叶-前壳核回路在抑制反应中的因果作用
摘要
停止不适当的反应需要前额叶-底丘脑超直接通路的参与。然而,前额-纹状体间接通路如何促成停止的知之甚少。在这项研究中,经颅超声刺激用于在纹状体中与任务相关的区域进行干预。功能性磁共振成像 (MRI) 显示在反应抑制期间壳核右前部的激活,并且对前壳核以及丘脑底核的超声刺激导致停止表现的显著损害。扩散成像进一步揭示了前壳核和下额叶皮层(IFC)右前部之间的显著结构连接,并且对前IFC的超声刺激也显示出显著受损的停止表现。这些结果表明,右前壳核和右前 IFC 对反应抑制有因果关系,并表明间接通路中的前 IFC-前壳核回路是反应抑制的重要途径。
图形摘要
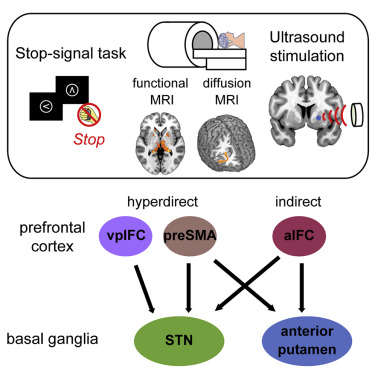
关键词
• 基底神经节
• 功能磁共振成像
• 神经影像学检查
• 无创脑刺激
• 反应抑制
• 停止信号任务
• 经颅超声刺激
• TUS
介绍
在特定情况下抑制已经开始但不适当的反应对于适应性行为至关重要。额叶-基底神经节相互作用,包括丘脑底核 (STN) 和纹状体,实现反应抑制。特别是,先前的神经影像学研究表明,连接下额叶皮层 (IFC)、辅助前运动区 (preSMA) 和右半球 STN 的超直接回路被认为是支持人类反应抑制的主要途径。神经系统疾病患者 STN 的深部脑刺激也证实了其重要性。
纹状体是直接和间接途径的主要门户。它接收来自额叶皮层的输入,并投射到全苍白球内部/外部或黑质。间接途径已经被提出作为停止的替代途径。先前的人类神经影像学研究报告了在反应抑制期间纹状体的激活。然而,人类纹状体对反应抑制的重要作用仍然很大程度上不为所知。
在本研究中,低强度经颅超声刺激(TUS)被用来对人类基底神经节进行干预。TUS可以无创地刺激大脑深层结构,空间精度高达几毫米。以前的研究报告称,TUS可以诱发短期或长期的抑制/增强活动,这取决于刺激方案。我们首先通过经颅磁刺激(TMS)测量运动诱发电位(MEP),证实了人类初级运动皮层(M1)的TUS后皮质活动的持续抑制(Stage 1;图1A )。随后,通过功能磁共振成像(fMRI)确定基底神经节中与任务相关的区域,以进行停止信号任务的反应抑制(Stage 2;图1A)。用TUS刺激纹状体中确定的区域,以检查反应抑制性能的破坏情况(Stage 3;图1A)。此外,大脑皮层区域被确定为纹状体的皮层对应区(Stage 4;图1B),并检查皮层区域的本质(Stage 5;图1B)。
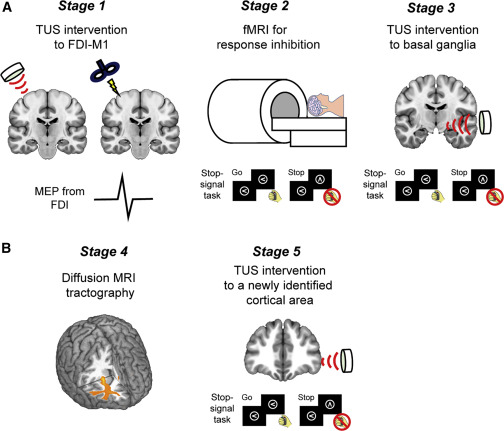
图1.实验设计概述
结果
通过TUS刺激M1区持续抑制皮质活性(Stage 1)
在TUS干预前后测量右侧**背侧骨间肌(FDI)的MEP,以验证TUS刺激后的持续抑制效果。使用fMRI测量大脑活动,同时受试者执行运动任务以识别每个受试者左半球M1(FDI-M1)中的FDI代表(图2A)。该任务旨在激活左半球.活化局限于中央沟前的区域(图2B)。每个受试者左半球的FDI-M1点都是TMS和TUS在在线导航系统的帮助下精准定位刺激的。
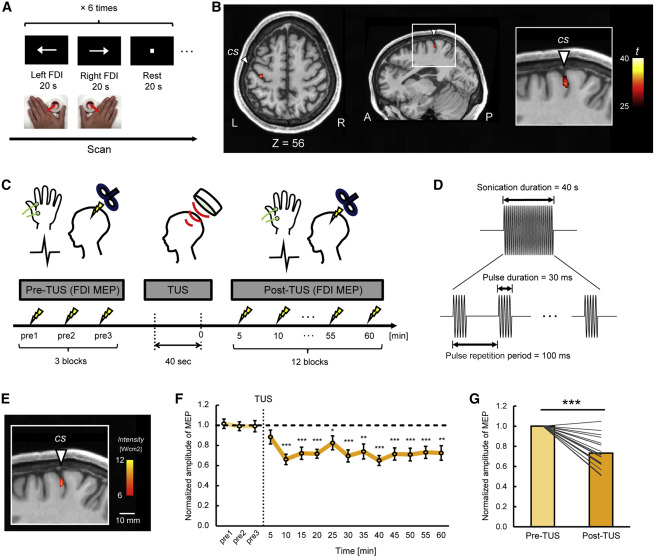
图2. FDI-M1识别和TUS干预(Stage 1)
通过针对已识别的FDI-M1点,在TUS刺激之前和之后测量来自右侧FDI肌肉的MEP(图2C)。通过发送单脉冲TMS刺激来记录MEP。使用四阵元换能器探头将一组40秒的脉冲超声(500 kHz)传送到左半球的目标FDI-M1,包括每100 ms一个30 ms的burst脉冲(图2D)。基于数值声学模拟的超声强度估计证实,FDI-M1目标区域被TUS有效靶向(图2E)。表S1总结了大脑中的峰值强度以及大脑和骨组织中的温度升高。图2F显示,MEP在TUS刺激后被抑制至少60分钟。与TUS刺激之前相比,TUS刺激后MEP显著下降(t(19) = 8.2,p = 1.1×10−7,配对 t 测试)(图 2G)。这些结果表明,TUS刺激诱导了皮质活性的持续抑制。
反应抑制期间的大脑激活(Stage 2)
受试者在 MRI 扫描仪内执行停止信号任务(图 3A)。使用停止信号反应时间 (SSRT) 评估行为效率。具有较短 SSRT 的受试者被认为在反应抑制方面更有效。行为数据总结在表 S2 中。在 IFC 中观察到反应抑制期间的显著激活,包括后 IFC(vpIFC 和 dpIFC)的腹侧和背侧部分,以及右半球的前 SMA(图 3B)。在皮层下区域,在 STN 和右半球纹状体中的壳核前部观察到任务相关的激活(图 3C)。这些结果与先前使用停止信号任务的研究一致。如先前报道的(图 3D),大脑皮层被分成 360 个功能区域,并且计算了分块激活以评估每个功能单元中的大脑活动(图 3E)。跨受试者的大脑活动和 SSRT 之间的相关性分析显示,vpIFC、dpIFC、前壳核和尾状核(图 S1A 和 S1B)具有很强的负相关性,这与之前的研究一致。
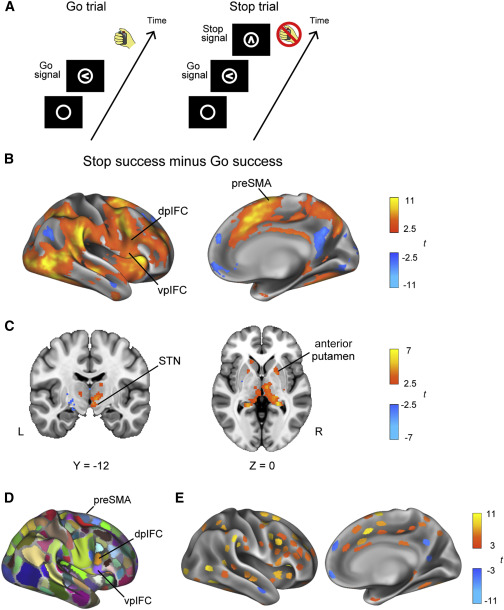
图3.反应抑制期间的停止信号任务和大脑活动(Stage 2)
我们确认独立比赛模型的三个假设在 fMRI 期间的行为数据集中得到满足:“go”成功试验的反应时间(RT)明显长于“stop”试验的反应时间(RT)。 ” 失败试验(t(29)= 10.4,p= 2.9 × 10-11,配对 t 检验)(图 S1C)。具有较长停止信号延迟 (SSD) 的“停止”故障试验的 RT 明显长于具有较短 SSD 的“停止”故障试验 (t(29) = 15.2, p = 2.4 × 10−15, 配对t 检验)(图 S1D)。图 S1E 显示了“停止”失败试验的比例与响应和停止过程的相对完成时间的函数关系图。这些图证实了“停止”失败试验的概率随着 SSD 的增加而增加。
TUS干预揭示了基底神经节对反应抑制的重要性(Stage 3)
我们检查了已确定的基底神经节区域在TUS干预下的反应抑制的本质(图4A;表S1),其中TUS刺激被应用于右半球的STN和前壳核(图4B)。作为对照,还调查了右半球壳核的后部,在那里没有观察到与任务有关的激活(图4B)。在TUS刺激前后(pre-and post-TUS),对停止信号任务的表现进行了监测,并进行了40s的超声刺激。
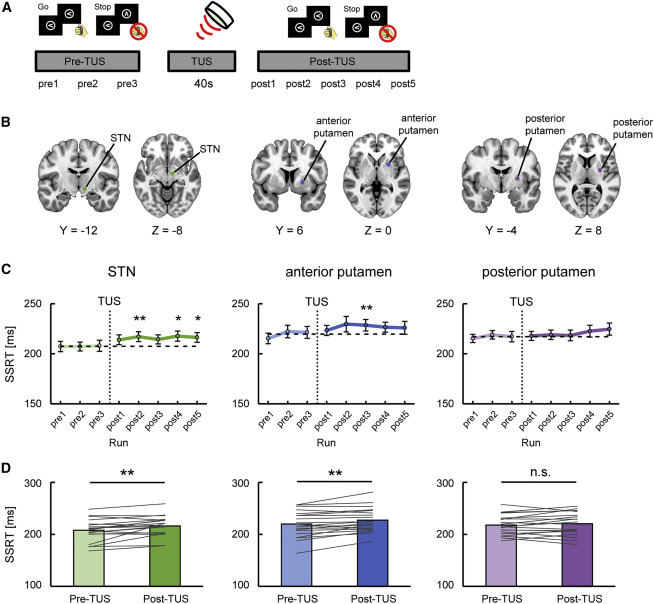
图4.TUS在反应抑制期间对基底神经节的影响(Stage 3)
图4C和4D显示了三个基底神经节区域在TUS刺激后的行为变化。STN在TUS后表现出反应抑制性能的持续破坏(平均差异。8.5毫秒,t(19)=3.4,p=3.4×10-3,配对t检验),证实了以前关于STN深部脑刺激的报告和STN的病变在神经系统疾病患者的结果。前壳核也显示了反应抑制性能的持续破坏(平均差异。7.3毫秒,t(19)=3.4,p=3.1×10-3,配对t检验)。这些显著性水平克服了Bonferroni校正的多重比较的3倍(受刺激区域的数量)。相比之下,后枕叶区没有表现出反应抑制性能的明显破坏(平均差异:2.8毫秒,t(19)=1.2,p=0.24,配对t检验)。
在 "go "的RT或 "go "和 "stop "的反应率(TUS前与TUS后)没有观察到明显的变化(图S2A、S2B、S2D和S2E)。刺激前壳核时,观察到SSD的显著变化(平均差异。7.4毫秒,t(19)=2.2,p=0.04,配对t检验)(图S2C),尽管经多重比较校正后差异不大。我们确认,在Stage 3的数据集中,独立比赛模型的三个假设得到了满足(图S2F-S2H)。
通过弥散性 MRI 确定基底神经节-脑皮质连接(Stage 4)
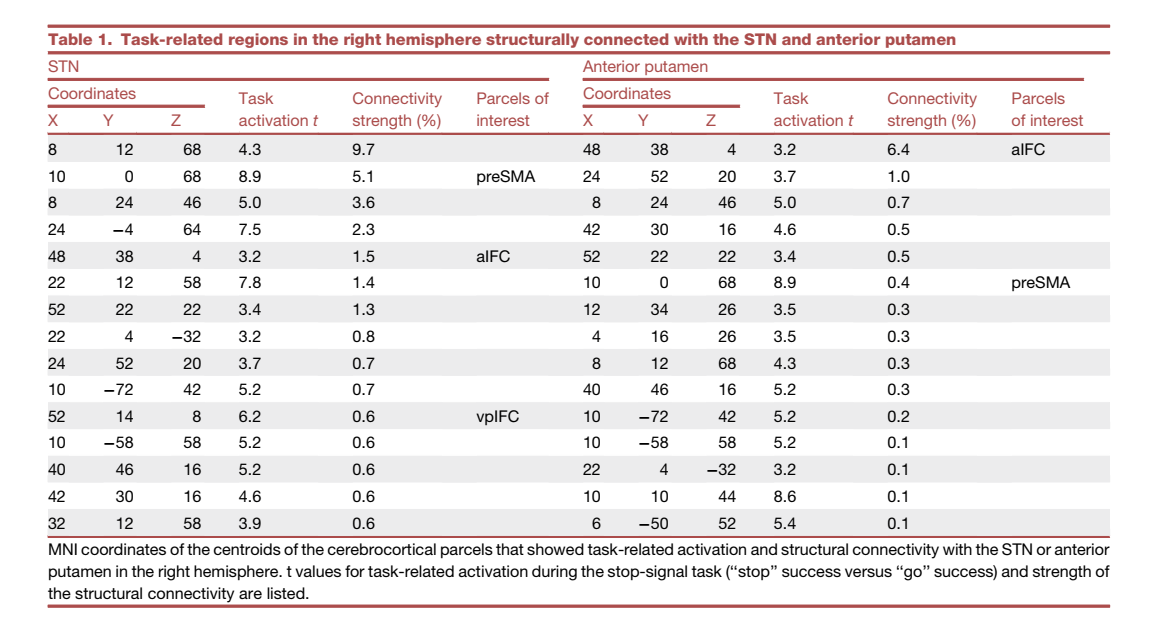
先前的研究已经使用弥散 MRI 揭示了 STN 和脑皮质区域之间的解剖学联系,并确定了 STN 及其网络参与反应抑制。我们接下来探索了已识别的基底神经节区域和大脑皮质区域之间的结构联系,以描绘反应抑制的间接途径。使用了来自人类连接组计划 (HCP) 的弥散 MRI 数据。右半球的 STN 被设置为种子,使用概率纤维束成像估计右半球大脑皮质区域的束(图 S3)。还检查了来自右半球前壳核的束(图 5A)。然后使用Stage 2的任务激活数据对纤维束成像数据进行阈值处理(图 5B)。表 1 显示了与基底神经节区域相连并且在反应抑制期间表现出任务激活的区域。如前所述(表 1),右半球的 vpIFC 和 preSMA 显示了与 STN 的任务相关激活和结构连接(Aron 等,2007;Forstmann 等,2012;Rae 等,2015)。另一方面,观察到右半球的前 IFC (aIFC) 与前壳核的结构连接性最强(图 5B 和 5C;表 1)。 aIFC 位于三角部的前端(图 S4)。
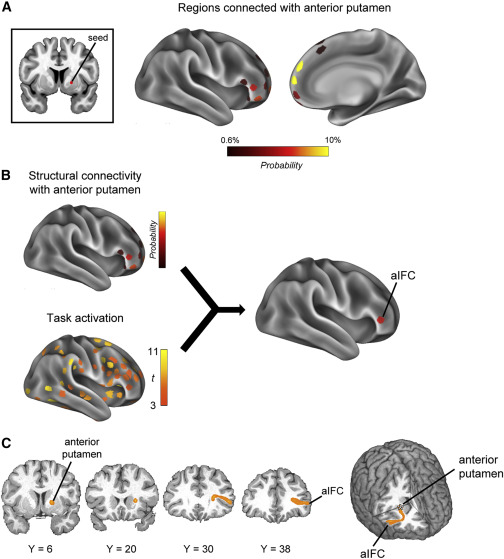
图5.前壳膜和脑皮质区域之间的结构连接(Stage 4)
表1右半球与STN和前壳体结构相连的任务相关区域
aIFC对于反应抑制至关重要(Stage 5)
我们进一步研究了已识别的 aIFC区 的重要性(图 6A;表 S1)。位于右半球的与 aIFC 相邻的中额叶皮层 (MFC) 也作为对照进行了研究,其中观察到了与任务相关的激活;然而,没有观察到与前壳核的强连接(图 6A)。图 6B 和 6C 显示了 TUS 刺激后的行为变化。在 TUS刺激 后,aIFC 表现出反应抑制性能的持续中断(平均差异:9.7 ms,t(19) = 4.9,p = 9.3 × 10-5,配对 t 检验)。这个显著性水平超过了多重比较的 Bonferroni 校正 2 倍(受刺激的皮质区域的数量)。相比之下,MFC 在 TUS 后没有表现出反应抑制性能的持续中断(平均差:2.1 ms,t(19) = 1.0,p = 0.35,配对 t 检验)。
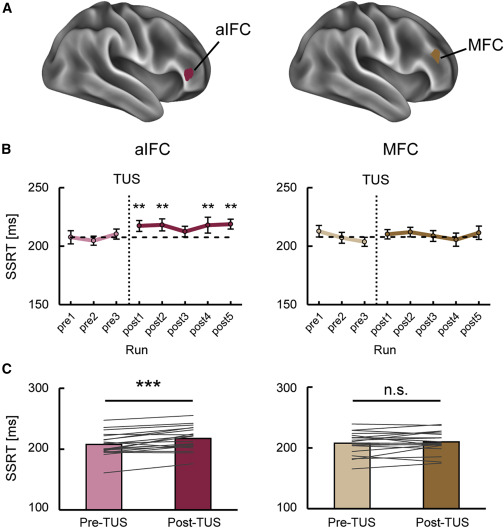
图6TUS在反应抑制期间对前额叶区域的影响(Stage 5)
在“go”RT,SSD或“go”和“stop”响应速率(TUS之前与之后)中基本上没有观察到显著变化(图S5)。然而,“go”RT在TUS之后延长(均差:9.6 ms,t(19) = 5.2,p = 4.8 ×10−5,配对t检验)和TUS后“停止”响应率下降(均差:1.3%,t(19)= 2.3,p = 0.031,配对t检验)(图S5B和S5E)。有趣的是,“go”RT在TUS后过程中显著增加,这是通过TUS后最后两次运行和前两次运行之间的差异来估计的(均值差:10.0 ms,t(19)= 3.2,p = 0.0045,配对t测试),而在TUS后过程中SSRT中没有观察到显著增加(均差:0.7 ms, t(19) = 0.2, p = 0.83, 配对 t 检验)。必须注意的是,“go”RT和“stop”响应速率的差异不能解释SSRT的差异。我们证实,独立比赛模型的三个假设在Stage 5的数据集中得到了满足(图S5F–S5H)。
讨论
本研究证明了纹状体对反应抑制的重要性。通过利用 TUS 的持续抑制效应,对右侧前壳核的刺激会导致停止表现受损。此外,使用弥散 MRI 确定了右半球前壳核和 aIFC 之间的结构连接。右侧 aIFC 的 TUS 刺激也导致停止表现受损。这些结果表明,间接途径中通过aIFC-前壳核回路的脑皮质-纹状体相互作用在反应抑制中起因果作用。
超声刺激的技术注意事项
本研究采用基于 Verhagen et al., 2019 年的 40 秒 TUS 协议的离线范式,采用该TUS协议在猕猴中观察到大脑连接的持续变化。本研究成功地证实了 FDI-M1 在stage 1 持续抑制皮质活动,持续至少 60 分钟。此外,在stage 3 观察到 TUS 中的行为效应与基底神经节(STN 和前壳核与后壳核)的分离。最近,一些研究报告了其他诱导持续兴奋性变化的方案,例如 15 分钟刺激(Zhang 等人., 2021) 和 80 秒 theta-burst 刺激 (Zeng et al., 2022)。通过 TUS 调节神经元活动被认为是由细胞膜的瞬时机械扰动引起的,从而改变了离子通道的通透性。然而,TUS 调制的精确神经机制需要进一步研究。
基底神经节在反应抑制中的作用
本研究基于Stage 2 明显的 fMRI 激活突出了纹状体中的右前壳核。观察到前壳核具有最强的 fMRI 激活,在纹状体中具有较大的空间范围。先前的研究也报道了前壳核的激活。值得注意的是,虽然峰值 t 值和空间范围没有前壳核中的那么大,但尾状核中似乎存在其他 fMRI 激活簇。几项研究提出,在反应抑制期间,右侧尾状核在间接通路中充当前额叶皮层的门户。因此,不能排除尾状核参与反应抑制的可能性。
对前壳核的干预导致stage 3停止表现的受损。据报道,纹状体中的病变会导致大鼠停止损害。纹状体已被提议作为间接途径的一部分参与反应抑制。先前的神经影像学研究报道纹状体的激活与提前停止的准备有关,表明纹状体通过前额-纹状体间接途径参与主动抑制控制。尽管在本研究的停止信号任务期间没有向受试者提供有助于准备停止反应的明确外部信息,但可以在停止信号任务的常见实验设置下采用主动抑制控制机制。
aIFC与基底神经节在反应抑制期间通过间接途径的相互作用
使用弥散MRI进行的结构连接分析显示,在stage 4,前壳核和aIFC之间有突出的连接。STN与preSMA的连接比与vpIFC的连接更强(表1),这与先前的报告一致。前壳核前区的情况也是如此,与preSMA的连接比与vpIFC的连接强(表1)。这些结果表明vpIFC对STN的特异性。
对aIFC的TUS刺激导致了在Stage 5停止时的SSRT(受损)。必须注意的是,"go"的RT也被延长了,而 "stop"的反应率在TUS到aIFC后被降低。“stop"反应率的降低不能解释SSRT的延长,因为SSRT是用整合法计算的,即使 "stop"反应率不完全是50%,SSRT也可以被精确估计。更关键的是,延长 "go"的RT不能解释延长的SSRT,因为放慢 "go"的RT会导致停止反应率增加,导致基于跟踪程序的SSD延长,这抵消了计算SSRT中的放慢成分。有趣的是,"go"的RT,但不是SSRT,在后TUS期间逐渐增加。SSRT被认为是由多种成分过程决定的,如注意力监测和停止本身。延长SSRT的一个可能解释是TUS刺激对aIFC造成的注意力监测受阻。另一种可能是,构成大脑皮层-纹状体间接通路的aIFC本身就能实现停止。还需要注意的是,aIFC属于似乎本身就能实现停止的神经元-大脑皮层网络。需要进一步的研究来区分aIFC和其他区域在反应抑制过程中的作用。
似乎值得总结一下本研究和以前研究中与反应抑制有关的右IFC区域。我们研究中的vpIFC和dpIFC(图S4A)基本对应于Aron及其同事(Aron等,2014)提出的区域,并基本对应于右额下回的操作区(图S4B)。中间IFC(mIFC)(图S4A),大脑活动非常弱,位于vp/dpIFC的前面,包括在右额下回的三角区(图S4B)。本研究中的aIFC(图S4A)位于mIFC的前面,部分包括在三角区中。其空间关系见图S4C。由于aIFC和vp/dpIFC被mIFC所干扰,而mIFC的大脑活动非常弱,所以aIFC可以被认为与Aron及其同事提出的区域不同(Aron等人,2014)。aIFC已被发现被激活(Cai等人,2014),但没有被强调。
本研究证明了 aIFC 前壳核回路的重要作用,并表明该回路在反应抑制期间充当间接通路的输入。 aIFC 还与 STN 具有结构性连接。因此,aIFC 可以通过分别与前壳核和 STN 相互作用,作为间接和超直接通路的共同输入(图 S6)。超直接通路被认为在反应抑制期间介导快速处理,而间接通路参与相对较慢的处理。因此,建议位于 vpIFC 前面的 aIFC 作为分层前额叶结构中的高级位点并协调快速超直接和慢速间接途径以实现反应抑制。
研究的局限性
本研究证实,在将TUS刺激应用于M1后,MEP的下降。然而,TUS诱导的兴奋性抑制可能不适用于更深层结构,例如基底神经节区域。需要进一步研究以澄清细胞水平上的确切活动变化。使用HCP数据(而不是本研究中受试者的数据)使用弥散MRI识别基底神经节和脑皮质区域之间的结构连接,这可能降低了分析的信噪比。
STAR★Methods
Method details
经颅超声刺激步骤(Stages 1, 3, and 5)
在Stages 1,一个四阵元环形阵列 TUS 换能器(NeuroFUS CTX-500,Brainbox Ltd,Cardiff,UK)与一个可编程射频放大器(Transducer Power Output System,TPO-203,Brainbox Ltd,Cardiff,UK)连接,使用的基频为 500 kHz,直径为 60 mm。放大器控制四个阵元的相位以调整超声刺激深度。超声刺激由每 100 毫秒重复一次的 30 毫秒burst脉冲组成,总超声刺激持续 40 秒(Verhagen 等,2019)。将水凝胶垫(Echo Gel Pad,Yasojima,Kobe,Japan)放置在换能器表面和受试者头皮之间,并在头皮-垫界面处应用超声凝胶(Aquasonic 100,Parker Laboratories,NJ,USA)。在线影像导航系统用于引导刺激识别出的目标区域。所用超声刺激在水中(即自由场)的空间峰值脉冲平均强度为 ISPPA = 30.0 W/cm2(空间峰值时间平均强度,ISPTA = 9.0 W/cm2)。颅骨的衰减降低了这些值。 k-Wave (Treeby and Cox, 2010) 是一个声学模拟工具箱,用于根据换能器位置计算每个受试者的声学焦点模拟。基于每个受试者的结构 MRI 估计皮肤、头皮、颅骨和脑组织的 3D 图,并用于模拟。为每个超声处理条件绘制超声处理后的强度场和温度升高。本研究中的受试者在 TUS 实验后没有报告任何不良反应。
在Stages 3和Stages 5,TUS 程序与Stages 1的相似。在Stages 3刺激右半球基底神经节的三个区域(STN、前壳核和后壳核),而在前额叶皮层中的两个区域受到刺激(aIFC 和 MFC)在Stages 5受到刺激。水中的强度为 ISPPA = 35.8 W/cm2 和 ISPTA = 10.7 W/cm2。目标的 MNI 坐标为:STN 为 (10, -12, -8),前壳核为 (20, 6, 0),后壳核为 (28, -4, 8),(48, 38 , 4) 为 aIFC 区域的质心, (44, 36, 22) 为 MFC 区域的质心。在第 3 阶段和第 5 阶段,受刺激区域在受试者之间被伪随机平衡。
At Stage 1, a four-element annular array TUS transducer (NeuroFUS CTX-500, Brainbox Ltd, Cardiff, UK), coupled with a programmable radiofrequency amplifier (Transducer Power Output System, TPO-203, Brainbox Ltd, Cardiff, UK), was used with a fundamental frequency of 500 kHz and diameter of 60 mm. The amplifier controlled the phasing of the four elements to adjust sonication depth. The ultrasound stimulation consisted of 30 ms bursts repeated every 100 ms, and the total sonication lasted for 40 s. Aqueous gel pads (Echo Gel Pad, Yasojima, Kobe, Japan) were placed between the surface of the transducer and the subject’s scalp, and ultrasound gel (Aquasonic 100, Parker Laboratories, NJ, USA) was applied at the scalp-pad interface. The online navigator was also used to stimulate the identified target region. The spatial-peak pulse-average intensity in water (free field) was ISPPA = 30.0 W/cm2 (for spatial-peak temporal-average intensity, ISPTA = 9.0 W/cm2). The attenuation by the skull lowers these values. k-Wave, an acoustic simulation toolbox, was used to calculate simulations of the acoustic focus for each subject based on transducer positions. 3D maps of the skin, scalp, skull, and brain tissues were estimated based on structural MRI of each subject and were used in the simulations. Intensity fields and temperature increase after sonication were mapped for each sonication condition. The subjects in this study reported no adverse effects after the TUS experiments.
At Stages 3 and 5, the TUS procedures were similar to those at Stage 1. Three regions in the basal ganglia (STN, anterior putamen, and posterior putamen) in the right hemisphere were stimulated at Stage 3, whereas two parcels in the prefrontal cortex (aIFC and MFC) in the right hemisphere were stimulated at Stage 5. Intensities in water were ISPPA = 35.8 W/cm2 and ISPTA = 10.7 W/cm2. The MNI coordinates of the targets were: (10, −12, −8) for the STN, (20, 6, 0) for the anterior putamen, (28, −4, 8) for the posterior putamen, (48, 38, 4) for the centroid of the aIFC parcel, and (44, 36, 22) for the centroid of the MFC parcel. The stimulated regions were pseudorandomly counterbalanced across subjects both at Stages 3 and 5.



;){kind=link}